









Rabbit p53 Polyclonal Antibody | anti-p53 antibody
Phospho-p53 (Ser392) Antibody
Predicted Reactivity: Pig (88%), Bovine (88%), Sheep (88%), Rabbit (88%)
Predicted Reactivity: Pig (88%), Bovine (88%), Sheep (88%), Rabbit (88%)
Tissue Specificity: Ubiquitous. Isoforms are expressed in a wide range of normal tissues but in a tissue-dependent manner. Isoform 2 is expressed in most normal tissues but is not detected in brain, lung, prostate, muscle, fetal brain, spinal cord and fetal liver. Isoform 3 is expressed in most normal tissues but is not detected in lung, spleen, testis, fetal brain, spinal cord and fetal liver. Isoform 7 is expressed in most normal tissues but is not detected in prostate, uterus, skeletal muscle and breast. Isoform 8 is detected only in colon, bone marrow, testis, fetal brain and intestine. Isoform 9 is expressed in most normal tissues but is not detected in brain, heart, lung, fetal liver, salivary gland, breast or intestine.
IF/ICC: 1:100-1:500
IHC: 1:50-1:200
Peptide ELISA: 1:20,000-1:40,000
Note: Interaction with BANP promotes nuclear localization (PubMed:15701641). Recruited into PML bodies together with CHEK2 (PubMed:12810724). Translocates to mitochondria upon oxidative stress (PubMed:22726440). Translocates to mitochondria in response to mitomycin C treatment (PubMed:27323408).Nucleus. Cytoplasm.
Note: Predominantly nuclear but localizes to the cytoplasm when expressed with isoform 4.Nucleus. Cytoplasm.
Note: Localized mainly in the nucleus with minor staining in the cytoplasm.Nucleus. Cytoplasm.
Note: Localized in the nucleus in most cells but found in the cytoplasm in some cells.Nucleus. Cytoplasm.
Note: Predominantly nuclear but translocates to the cytoplasm following cell stress.Nucleus. Cytoplasm.
Note: Localized mainly in the nucleus with minor staining in the cytoplasm.Nucleus. Cytoplasm.
Note: Localized in both nucleus and cytoplasm in most cells. In some cells, forms foci in the nucleus that are different from nucleoli.Cytoplasm.
Application Data
(At 25 degree C. The primary antibody was diluted at 1/200 and incubated with the sample for 1 hour at 37 degree C. An Alexa Fluor 594 conjugated goat anti-rabbit IgG (H+L) Ab, diluted at 1/600, was used as the secondary antibody.)
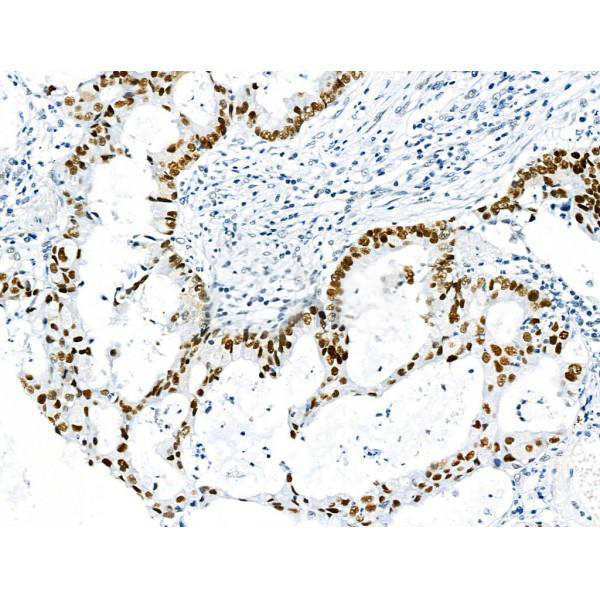
IHC (Immunohistochemistry)
(At 1/100 staining Human gastric cancer by IHC-P. The sample was formaldehyde fixed and a heat mediated antigen retrieval step in citrate buffer was performed. The sample was then blocked and incubated with the primary antibody at 4 degree C overnight. An HRP conjugated anti-Rabbit antibody was used as the secondary antibody.)
IHC (Immunohistchemistry)
(At 1/100 staining Human gastric cancer by IHC-P. The sample was formaldehyde fixed and a heat mediated antigen retrieval step in citrate buffer was performed. The sample was then blocked and incubated with the primary antibody at 4 degree C overnight. An HRP conjugated anti-Rabbit antibody was used as the secondary antibody.)
IHC (Immunohistochemistry)
(At 1/100 staining Human esophageal cancer by IHC-P. The sample was formaldehyde fixed and a heat mediated antigen retrieval step in citrate buffer was performed. The sample was then blocked and incubated with the primary antibody at 4 degree C overnight. An HRP conjugated anti-Rabbit antibody was used as the secondary antibody.)
IHC (Immunohistochemistry)
(At 1/100 staining Human kidney cancer and adjacent normal tissues by IHC-P. The sample was formaldehyde fixed and a heat mediated antigen retrieval step in citrate buffer was performed. The sample was then blocked and incubated with the primary antibody at 4 degree C overnight. An HRP conjugated anti-Rabbit antibody was used as the secondary antibody.)
IHC (Immunohistchemistry)
(At 1/100 staining Human ovarian cancer and adjacent normal tissues by IHC-P. The sample was formaldehyde fixed and a heat mediated antigen retrieval step in citrate buffer was performed. The sample was then blocked and incubated with the primary antibody at 4 degree C overnight. An HRP conjugated anti-Rabbit antibody was used as the secondary antibody.)
IHC (Immunohistochemistry)
(At 1/100 staining Mouse brain tissue by IHC-P. The sample was formaldehyde fixed and a heat mediated antigen retrieval step in citrate buffer was performed. The sample was then blocked and incubated with the primary antibody at 4 degree C overnight. An HRP conjugated anti-Rabbit antibody was used as the secondary antibody.)
IHC (Immunohistochemistry)
(At 1/100 staining Human lung cancer by IHC-P. The sample was formaldehyde fixed and a heat mediated antigen retrieval step in citrate buffer was performed. The sample was then blocked and incubated with the primary antibody at 4 degree C overnight. An HRP conjugated anti-Rabbit antibody was used as the secondary antibody.)
IHC (Immunohistochemistry)
(At 1/100 staining Human lung cancer by IHC-P. The sample was formaldehyde fixed and a heat mediated antigen retrieval step in citrate buffer was performed. The sample was then blocked and incubated with the primary antibody at 4 degree C overnight. An HRP conjugated anti-Rabbit antibody was used as the secondary antibody.)
WB (Western Blot)
(Western blot analysis of p53 (Phospho-Ser392) using nocodazole treated HeLa whole cell lysates.-/+ means absence or presence of N peptide(non-phospho peptide) and P peptide(phospho peptide).)
WB (Western Blot)
(Western blot analysis of extracts from 293, using p53 (Phospho-Ser392) Antibody.)


















