Understanding Antibody Glycosylation, Its Types, Importance & More
 Cynthia
11 min read
Cynthia
11 min read
In this Article
All of the products listed in AAA Biotech’s catalog are strictly for research-use only (RUO).
Biological medicines, particularly monoclonal
antibodies (mAbs), represent one of the fastest-growing and most
revolutionary classes of biomedical research, offering lifelines against chronic
diseases, autoimmune disorders, and cancer.
These molecules are engineered marvels, yet their clinical efficacy, safety profile, and stability do not depend solely on their genetic sequence. Instead, their function hinges entirely on tiny, often-overlooked sugar chains—glycans—covalently attached to them.
If the biopharmaceutical industry fails to precisely control these carbohydrates, the most advanced drugs developed can become less potent, unpredictable, or even toxic.
Glycosylation is not mere decoration; it is the functional operating system of the protein, and precise control over the resulting structure is now recognized as a Critical Quality Attribute (CQA) in modern biomanufacturing.
What You Will Learn in This Web Blog
You will learn the fundamental nature of glycosylation in biology, differentiating between N-linked glycans and O-linked glycans, and understanding what glycosylated proteins actually are. Crucially, we will detail how glycosylation of antibody structures dictates immune function, providing deep insight into the importance of protein glycosylation in health, disease, and the development of revolutionary therapeutic monoclonal antibodies.
The Biological Imperative: What is Glycosylation and Why is it Important?
Glycosylation is defined as one of the most common, ubiquitous, and complex
forms of post-translational modification (PTM), involving the covalent addition
of sugar moieties to specific amino acids. This intricate biochemical process is
essential for the production of fully functional glycoproteins, glycolipids, and
other glycoconjugates.
The Biological Scale and Mechanism of Glycosylation
- Approximately half of all proteins typically expressed in a cell, and
possibly over 70% of the eukaryotic proteome, undergo glycosylation.
- Glycosylation occurs primarily within the biosynthetic-secretory pathway,
mainly in the Endoplasmic Reticulum (ER) and Golgi apparatus.
- The synthesis of complex sugar structures involved in glycosylation is not
template-driven like DNA or protein synthesis.
- Instead, the process relies on the availability and specificity of enzymes
called glycosyltransferases (Gtfs) and glycosidases.
- This process is sequential and ordered, tightly regulated by the cell
through the trafficking of proteins across distinct Golgi cisternae, each with
unique enzyme concentrations.
- Because the outcome depends on cellular environment rather than a strict
genetic template, the resulting glycan structures - collectively called the
"glycome"- exhibit immense structural diversity, far surpassing that of the
proteome.
- This large heterogeneity in glycan structures allows the body to produce
complex, environmentally responsive molecular signals.
- The glycan profile effectively acts as a real-time snapshot of an organism's
physiological state, reflecting inflammation, disease, or other cellular
conditions.
Deciphering the Glycome: Understanding the Types of Glycosylation
The incredible diversity of functions carried out by what are glycosylated
proteins stems directly from the varied types of Glycosylation possible,
distinguished by the chemical linkage between the sugar and the protein.
A. N-Linked Glycans: The Structural Powerhouse
Mechanism of N-Glycosylation
N-linked glycosylation involves the attachment of an oligosaccharide (or glycan) to the nitrogen atom (the amide nitrogen) of the side chain of an Asparagine (Asn) residue. This linkage typically occurs within a consensus amino acid sequence, Asn-X-Ser or Asn-X-Thr (where X can be any amino acid except proline).
The process that defines N-glycosylation is primarily co-translational, meaning it occurs as the protein is being synthesized in the ER lumen. It is a complex process initiated by the building of a large precursor glycan structure which is temporarily attached to a specialized lipid called dolichol phosphate. This pre-assembled oligosaccharide is then transferred en bloc to the nascent protein.
Maturation and Diversity
Once transferred, the protein moves to the Golgi apparatus, where glycan processing, or terminal glycosylation, begins. This involves the precise trimming of some sugars (catalyzed by glycosidases) and the addition of others (catalyzed by Gtfs). This step-wise maturation pathway, segregated across distinct Golgi cisternae, generates highly diverse oligosaccharides. This processing results in three major mature structures:
- High-Mannose Glycans: Structures that largely retain the
mannose core and escape full terminal processing.
- Hybrid N-Glycans: Structures that exhibit characteristics
of both high-mannose and complex types.
- Complex N-Glycans: Highly diversified structures containing
various combinations of fucose, sialic acid (N-acetylneuraminic acid),
galactose, and N-acetylglucosamine.
B. O-Linked Glycans: The Signaling Gatekeeper
Mechanism and Location
In contrast to the N-linked mechanism, O-linked glycans attach a sugar moiety to the hydroxyl oxygen of either Serine (Ser) or Threonine (Thr) residues. While less common, linkages can also occur on Tyrosine, hydroxylysine, or hydroxyproline side chains.
O-linked glycosylation at its core is:
- O-linked glycosylation is primarily a post-translational process, occurring
after the protein is fully synthesized and folded.
- It does not utilize a large lipid-linked precursor; instead, sugars are
added stepwise.
- The process commonly begins with the addition of N-acetylgalactosamine (GalNAc) for mucin-type O-glycosylation.
- Localization differs from N-linked glycosylation.
- N-linked glycosylation starts in the ER lumen.
- O-linked glycosylation can occur in the ER, Golgi apparatus, cytosol, and
nucleus.
- In the nucleus and cytoplasm, O-GlcNAc modification regulates gene
expression and signal transduction.
- O-GlcNAc often competes with phosphorylation for the same residue
sites.
- N-linked glycosylation, being co-translational, is critical for structural
integrity during protein synthesis.
- O-linked modification, occurring post-synthesis, acts as a dynamic
regulatory marker on mature proteins.
C. Other Types of Glycosylation
While N-linked and O-linked are the most prevalent in mammals, other types exist, including:
- C-linked Glycans: A rare form where mannose binds to a
carbon on a tryptophan residue, usually in the W-X-X-W sequence.
- Phosphoglycosylation: Where a glycan binds to serine via a
phosphodiester bond.
- Glypiation: The addition of a GPI anchor, linking proteins
to lipids.
Table 01:Comparison of Major Glycosylation Types
| Aspect | N-Linked Glycosylation | O-Linked Glycosylation |
|---|---|---|
| Attachment Site | Amide nitrogen of Asparagine (Asn) | Hydroxyl oxygen of Serine (Ser) or Threonine (Thr) |
| Linkage Type | beta-1,4 glycosidic bond | alpha-O-glycosidic bond (often) |
| Timing | Primarily Co-translational (during synthesis) | Primarily Post-translational (after synthesis/folding) |
| Precursor Requirement | Lipid-linked oligosaccharide (Dolichol phosphate) | Direct transfer from sugar nucleotides (no lipid precursor) |
| Core Structure | Complex precursor - transferred en bloc | Single sugar initiation (e.g., GalNAc or GlcNAc) |
| Primary Biological Role | Folding, stability, protein localization/secretion, and immune response | Cell-cell interaction, signal transduction, expression/stability |
The Biological Mandate: What Does Glycosylation Do?
The detailed examination of what glycosylation does reveals its essential role
as a structural, communication, and regulatory modification imperative for
protein functionality, stability, and interaction within the cellular
environment.
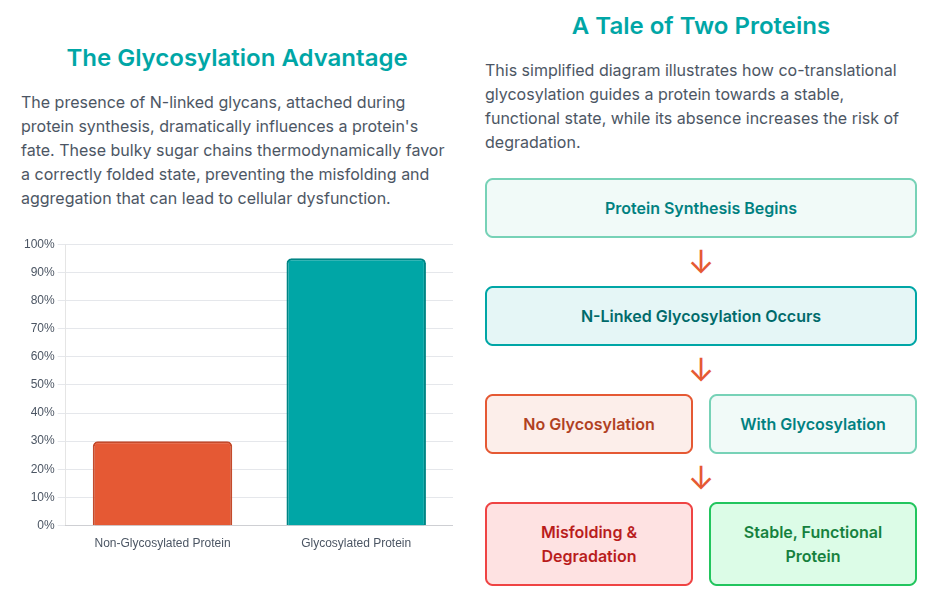
Research has shown that N-linked glycans actively help proteins fold, and
the removal of these sugar chains alters protein folding kinetics and stability.
The PTM process is critical as well in triggering a immune response. The mechanism can be easily digested via the following infographic:
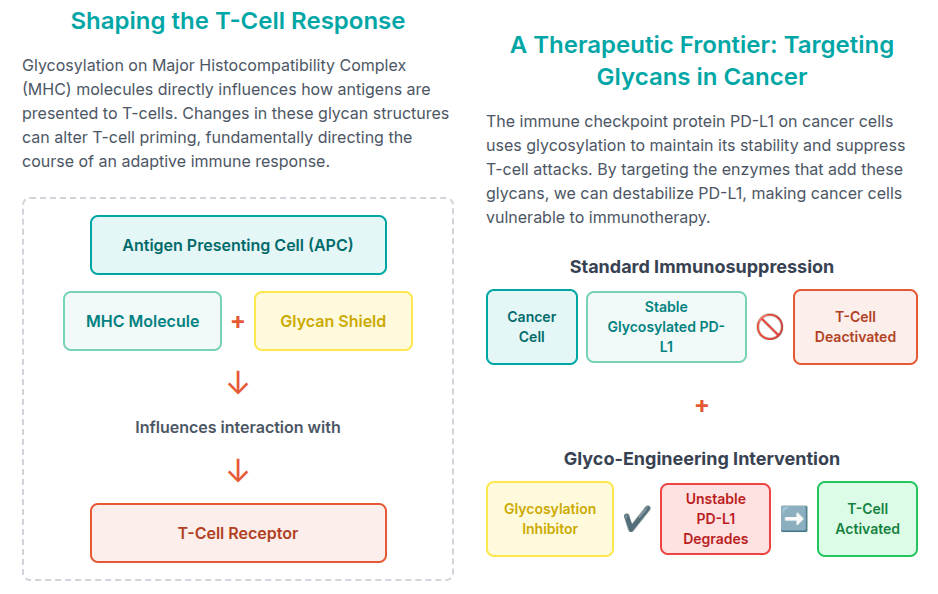
The Sweet Core of Immunity: Glycosylation and Antibodies (A Detailed Look)
Immunoglobulin G (IgG) antibodies are the archetypal glycoproteins. For therapeutic applications, understanding the precise glycosylation antibody profile is paramount, as the sugar chains fundamentally dictate the efficacy of the drug.
IgG Structure and Glycosylation Sites
IgG antibodies are Y-shaped molecules composed of two heavy chains and two light chains. The antibody is functionally divided into two main sections:
- Fragment variable (Fab): Contains the antigen-binding sites
formed by the hypervariable regions.
- Fragment crystallizable (Fc): Mediates interactions with
immune effector cells and complement factors.
The Conserved Fc Glycan (N297)
The most critical and universally conserved modification is the N-linked
glycosylation site located at Asparagine
297 (N297) within the CH2 domain of the Fc region.
This site is characterized by an occupancy that is virtually 100% in circulating IgG, confirming its essential nature. The N297 glycan is structurally vital: it fills a large pocket located between the two CH2 domains, maintaining the overall conformation and correct distance between the heavy chains.
The N297 glycan is typically a complex-type biantennary structure with several key variable extensions:
- Core Fucose (Fuc): Attached to the innermost GlcNAc.
- Galactose (Gal): Attached to the terminal GlcNAc
branches.
- Bisecting GlcNAc: Attached to the core mannose.
- Sialic Acid (Neu5Ac): The terminal cap, attached to
galactose.
Non-Conserved Fab Glycans and O-Linked Sites
While the Fc glycan provides the structural scaffolding, additional N-linked
sites are sometimes found in the Fab portion, often emerging near the
antigen-binding regions (hypervariable loops).
- These Fab glycans act as local modulators, sometimes enhancing
antigen-binding affinity by altering local flexibility or charge
distribution.
- This distinction - conserved Fc glycosylation for systemic immune control
and variable Fab glycosylation for fine-tuned antigen interaction - adds a new
dimension to antibody diversification beyond gene recombination.
- Certain antibody subclasses, such as IgG3, also carry O-linked glycans in
their extended hinge regions, and sometimes up to three O-linked sites per heavy
chain.
- The O-linked glycans are thought to protect against proteolytic cleavage and
maintain the flexible hinge conformation crucial for effective antigen
engagement and immune signaling.
The Functional Impact: How Glycans Drive Antibody Action
The importance of protein glycosylation in antibodies is best demonstrated by
how changes to the conserved N297 glycan profile determine the intensity and
manner of the resulting immune response, primarily by mediating binding to Fc
gamma receptors on immune cells.
Controlling Effector Functions
The Fc region initiates critical effector functions such as Antibody-Dependent Cell-mediated Cytotoxicity (ADCC), Antibody-Dependent Cellular Phagocytosis (ADCP), and Complement-Dependent Cytotoxicity (CDC).
The Fc glycan acts as a key regulatory element, utilizing both steric and allosteric mechanisms to shift the balance of Type I and Type II engagement. The specific structure of the N297 glycan dictates the precise conformation of the CH2 domain, thereby determining the accessibility and affinity for various effector cell receptors.
Fucosylation: The ADCC Switch
Core fucosylation is the addition of fucose to the innermost GlcNAc residue of the N297 glycan. It is perhaps the most functionally significant modification in therapeutic antibody design.
- Mechanism: Core fucosylation has been shown to exert a
strong adverse effect on the binding affinity for CD16a. This receptor is
predominantly expressed on Natural Killer (NK) cells, the primary drivers of
ADCC.
- Result: When the core fucose is removed, the resulting
low-fucosylated (or afucosylated) variants show substantially enhanced affinity.
This structural change translates into a vastly improved ability to recruit NK
cells, enhancing ADCC activity dramatically - often by 10-fold or more.
This makes fucosylation a highly sought-after characteristic for
oncology mAbs
designed to kill tumor cells.
Galactosylation and Sialylation: The Anti-Inflammatory Modulators
Galactose (Gal) and Sialic Acid (Neu5Ac) represent the final terminal residues added to the N297 glycan branches and serve as indicators of the inflammatory state.
- Sialylation: The presence of terminal sialic acid residues
is strongly associated with an anti-inflammatory function, often observed in
pooled therapeutic intravenous immunoglobulin (IVIg). However, the functional
impact is complex; studies indicate that sialylation, when present alongside
core fucosylation, results in a noticeable decrease in ADCC activity.
- Galactosylation and Disease: These terminal sugars are intimately linked to immune health. Analysis of IgG from patients with active Juvenile Idiopathic Arthritis (JIA), for example, demonstrated a robust reduction in both galactosylation and sialylation compared to healthy controls. This functional consequence is directly observed in receptor binding, as the interaction of IgG with Fc receptor-like protein 5 (FcRL5) was found to be mediated specifically through both galactose and sialic acid residues.
Table 02: Key Glycan Modifications and Their Impact
on IgG Function
| Glycan Modification | Location on N297 Glycan | Functional Consequence | Biomedical Implication (What Does Glycosylation Do?) |
|---|---|---|---|
| Core Fucose (Fuc) | Attached to the core GlcNAc | Dramatically reduces affinity for activating CD16a (NK cells) | Target for removal (Afucosylation) to enhance therapeutic killing (ADCC) |
| Galactose (Gal) | Attached to terminal GlcNAc branches | Required for full anti-inflammatory potential; Influences CDC activity | Reduction (Agalactosylation) is a biomarker for severe autoimmune disease (e.g., RA) |
| Sialic Acid (Neu5Ac) | Terminal cap on Gal residues | Strong anti-inflammatory signal; Associated with reduced disease activity | High levels are desired for anti-inflammatory therapies (e.g., IVIg) |
| Bisecting GlcNAc | Attached to the Man core | Generally enhances CD16a binding and ADCC | Modulation strategy for optimized next-generation mAbs |
Conclusion: The Future is Sweet
When we ask, “What is glycosylation when it comes to biology?”, the answer is
simple yet profound: it’s the sugar code that shapes molecular life. From
determining how an antibody binds to a virus to how long it persists in the
body, glycosylation governs immune structure, immune stability, and immune
performance. Understanding antibody-related glycosylation mechanisms and types
of glycans not only deepens our insight into immunity but also advances the
design of life-changing biotherapies.
As analytical methods continue to improve, advancing glycomics research ensures that the sophisticated language of sugar chains will translate directly into safer treatments and superior patient outcomes.
Faq's
How does carbohydrate attachment modify monoclonal antibodies (mAbs)?
Attachment of glycans stabilizes mAbs, influences their three-dimensional structure, and modulates their interactions with immune cells. These changes directly affect therapeutic potency, safety, and function, making glycosylation a critical attribute for mAb drugs.
What functional role do glycan structures play in immunological processes?
Glycan patterns regulate antibody effector functions like ADCC and CDC, control immune recognition, and can tune an antibody's response toward inflammation or immune suppression. They help signal the body's immune state, impacting disease progression or therapy.
Where specifically are the sugar molecules covalently linked on antibody structures (IgGs)?
The most conserved glycosylation site is at asparagine 297 (N297) in the CH2 domain of the Fc region. Additional glycans can modify Fab regions near antigen-binding sites, and some IgG subclasses (like IgG3) include O-linked sugars in their hinge region.
What biological consequences do the addition of sugar chains cause for an antibody molecule?
Sugar chains improve antibody stability, prevent aggregation, and modulate binding to Fc receptors. They dictate how effectively an antibody can trigger immune cell responses and clearance rates in therapies.
Why is precise glycan profiling a necessary step in the development and quality control of therapeutic biopharmaceuticals?
Accurate glycan profiling ensures product consistency, safety, and efficacy, minimizing batch-to-batch variation and adverse immune reactions. Biomanufacturers rely on glycoengineering and analytical techniques to maintain and verify these critical quality attributes.
Why are decreased levels of galactosylation and sialylation frequently observed in chronic autoimmune diseases?
Reduced galactosylation and sialylation are associated with a pro-inflammatory immune profile; these changes have been documented in conditions like rheumatoid arthritis and juvenile idiopathic arthritis. They indicate heightened disease activity and help guide both diagnosis & therapeutic development.
Lead Clinical Research Coordinator (LCRC)
Cynthia Lee is the President of AAA Biotech and specializes in understanding highly validated and characterized monoclonal/polyclonal antibodies, recombinant proteins, and ELISA kits.
Related Posts