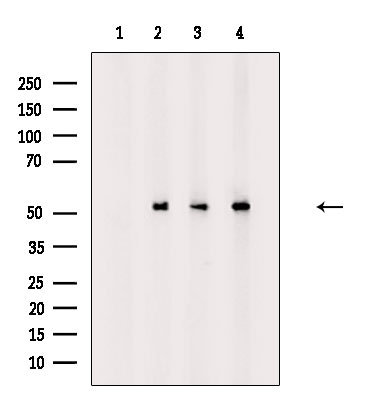
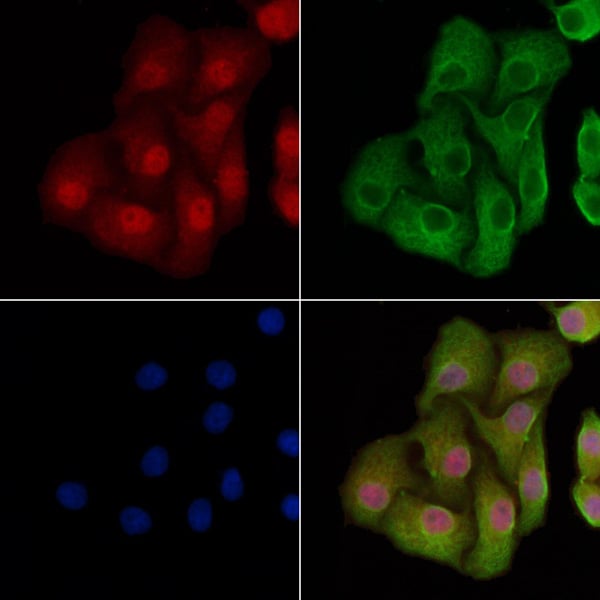
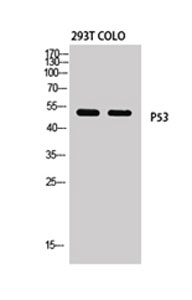
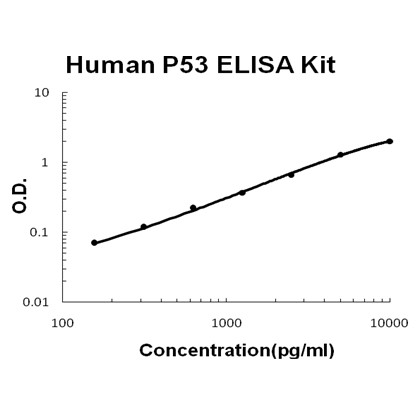
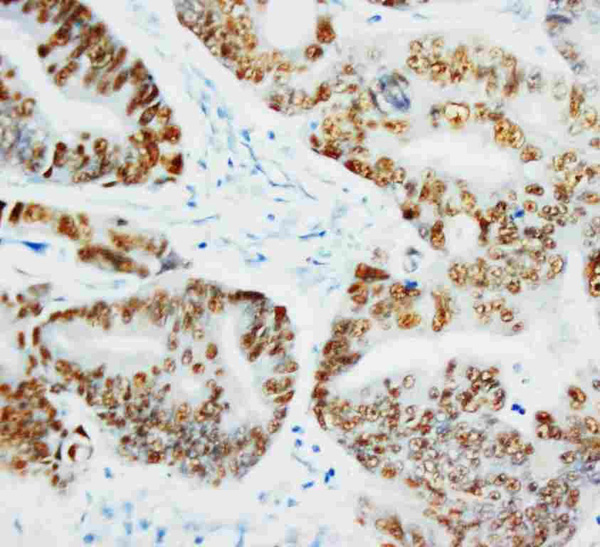
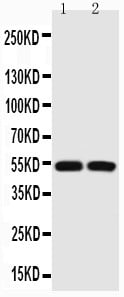
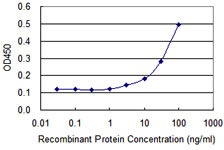
Applicable Applications for anti-TP53 antibody
ICC (Immunocytochemistry), IF (Immunofluorescence), WB (Western Blot)
Immunogen
A synthesized peptide derived from human p53.
Similarity
The nuclear export signal acts as a transcriptional repression domain. The TADI and TADII motifs (residues 17 to 25 and 48 to 56) correspond both to 9aaTAD motifs which are transactivation domains present in a large number of yeast and animal transcription factors. Belongs to the p53 family.
Post Translational Modifications
Acetylated. Acetylation of Lys-382 by CREBBP enhances transcriptional activity. Deacetylation of Lys-382 by SIRT1 impairs its ability to induce proapoptotic program and modulate cell senescence. Deacetylation by SIRT2 impairs its ability to induce transcription activation in a AKT-dependent manner.Phosphorylation on Ser residues mediates transcriptional activation. Phosphorylated by HIPK1 (By similarity). Phosphorylation at Ser-9 by HIPK4 increases repression activity on BIRC5 promoter. Phosphorylated on Thr-18 by VRK1. Phosphorylated on Ser-20 by CHEK2 in response to DNA damage, which prevents ubiquitination by MDM2. Phosphorylated on Ser-20 by PLK3 in response to reactive oxygen species (ROS), promoting p53/TP53-mediated apoptosis. Phosphorylated on Thr-55 by TAF1, which promotes MDM2-mediated degradation. Phosphorylated on Ser-33 by CDK7 in a CAK complex in response to DNA damage. Phosphorylated on Ser-46 by HIPK2 upon UV irradiation. Phosphorylation on Ser-46 is required for acetylation by CREBBP. Phosphorylated on Ser-392 following UV but not gamma irradiation. Phosphorylated on Ser-15 upon ultraviolet irradiation; which is enhanced by interaction with BANP. Phosphorylated by NUAK1 at Ser-15 and Ser-392; was initially thought to be mediated by STK11/LKB1 but it was later shown that it is indirect and that STK11/LKB1-dependent phosphorylation is probably mediated by downstream NUAK1. It is unclear whether AMP directly mediates phosphorylation at Ser-15. Phosphorylated on Thr-18 by isoform 1 and isoform 2 of VRK2. Phosphorylation on Thr-18 by isoform 2 of VRK2 results in a reduction in ubiquitination by MDM2 and an increase in acetylation by EP300. Stabilized by CDK5-mediated phosphorylation in response to genotoxic and oxidative stresses at Ser-15, Ser-33 and Ser-46, leading to accumulation of p53/TP53, particularly in the nucleus, thus inducing the transactivation of p53/TP53 target genes. Phosphorylated by DYRK2 at Ser-46 in response to genotoxic stress. Phosphorylated at Ser-315 and Ser-392 by CDK2 in response to DNA-damage. Phosphorylation at Ser-15 is required for interaction with DDX3X and gamma-tubulin.Dephosphorylated by PP2A-PPP2R5C holoenzyme at Thr-55. SV40 small T antigen inhibits the dephosphorylation by the AC form of PP2A.May be O-glycosylated in the C-terminal basic region. Studied in EB-1 cell line.Ubiquitinated by MDM2 and SYVN1, which leads to proteasomal degradation. Ubiquitinated by RFWD3, which works in cooperation with MDM2 and may catalyze the formation of short polyubiquitin chains on p53/TP53 that are not targeted to the proteasome. Ubiquitinated by MKRN1 at Lys-291 and Lys-292, which leads to proteasomal degradation. Deubiquitinated by USP10, leading to its stabilization. Ubiquitinated by TRIM24, RFFL, RNF34 and RNF125, which leads to proteasomal degradation. Ubiquitination by TOPORS induces degradation. Deubiquitination by USP7, leading to stabilization. Isoform 4 is monoubiquitinated in an MDM2-independent manner. Ubiquitinated by COP1, which leads to proteasomal degradation. Ubiquitination and subsequent proteasomal degradation is negatively regulated by CCAR2. Polyubiquitinated by C10orf90/FATS, polyubiquitination is 'Lys-48'-linkage independent and non-proteolytic, leading to TP53 stabilization (By similarity).Monomethylated at Lys-372 by SETD7, leading to stabilization and increased transcriptional activation. Monomethylated at Lys-370 by SMYD2, leading to decreased DNA-binding activity and subsequent transcriptional regulation activity. Lys-372 monomethylation prevents interaction with SMYD2 and subsequent monomethylation at Lys-370. Dimethylated at Lys-373 by EHMT1 and EHMT2. Monomethylated at Lys-382 by KMT5A, promoting interaction with L3MBTL1 and leading to repress transcriptional activity. Dimethylation at Lys-370 and Lys-382 diminishes p53 ubiquitination, through stabilizing association with the methyl reader PHF20. Demethylation of dimethylated Lys-370 by KDM1A prevents interaction with TP53BP1 and represses TP53-mediated transcriptional activation. Monomethylated at Arg-333 and dimethylated at Arg-335 and Arg-337 by PRMT5; methylation is increased after DNA damage and might possibly affect TP53 target gene specificity.Sumoylated with SUMO1. Sumoylated at Lys-386 by UBC9.
Subunit Structure
Forms homodimers and homotetramers. Binds DNA as a homotetramer. Interacts with AXIN1. Probably part of a complex consisting of TP53, HIPK2 and AXIN1 (By similarity). Interacts with histone acetyltransferases EP300 and methyltransferases HRMT1L2 and CARM1, and recruits them to promoters. Interacts (via C-terminus) with TAF1; when TAF1 is part of the TFIID complex. Interacts with ING4; this interaction may be indirect. Found in a complex with CABLES1 and TP73. Interacts with HIPK1, HIPK2, and TP53INP1. Interacts with WWOX. May interact with HCV core protein. Interacts with USP7 and SYVN1. Interacts with HSP90AB1. Interacts with CHD8; leading to recruit histone H1 and prevent transactivation activity (By similarity). Interacts with ARMC10, BANP, CDKN2AIP, NUAK1, STK11/LKB1, UHRF2 and E4F1. Interacts with YWHAZ; the interaction enhances TP53 transcriptional activity. Phosphorylation of YWHAZ on 'Ser-58' inhibits this interaction. Interacts (via DNA-binding domain) with MAML1 (via N-terminus). Interacts with MKRN1. Interacts with PML (via C-terminus). Interacts with MDM2; leading to ubiquitination and proteasomal degradation of TP53. Directly interacts with FBXO42; leading to ubiquitination and degradation of TP53. Interacts (phosphorylated at Ser-15 by ATM) with the phosphatase PP2A-PPP2R5C holoenzyme; regulates stress-induced TP53-dependent inhibition of cell proliferation. Interacts with PPP2R2A. Interacts with AURKA, DAXX, BRD7 and TRIM24. Interacts (when monomethylated at Lys-382) with L3MBTL1. Isoform 1 interacts with isoform 2 and with isoform 4. Interacts with GRK5. Binds to the CAK complex (CDK7, cyclin H and MAT1) in response to DNA damage. Interacts with CDK5 in neurons. Interacts with AURKB, SETD2, UHRF2 and NOC2L. Interacts (via N-terminus) with PTK2/FAK1; this promotes ubiquitination by MDM2. Interacts with PTK2B/PYK2; this promotes ubiquitination by MDM2. Interacts with PRKCG. Interacts with PPIF; the association implicates preferentially tetrameric TP53, is induced by oxidative stress and is impaired by cyclosporin A (CsA). Interacts with SNAI1; the interaction induces SNAI1 degradation via MDM2-mediated ubiquitination and inhibits SNAI1-induced cell invasion. Interacts with KAT6A. Interacts with UBC9. Interacts with ZNF385B; the interaction is direct. Interacts (via DNA-binding domain) with ZNF385A; the interaction is direct and enhances p53/TP53 transactivation functions on cell-cycle arrest target genes, resulting in growth arrest. Interacts with ANKRD2. Interacts with RFFL and RNF34; involved in p53/TP53 ubiquitination. Interacts with MTA1 and COP1. Interacts with CCAR2 (via N-terminus). Interacts with MORC3. Interacts (via C-terminus) with POU4F2 isoform 1 (via C-terminus). Interacts (via oligomerization region) with NOP53; the interaction is direct and may prevent the MDM2-mediated proteasomal degradation of TP53. Interacts with AFG1L; mediates mitochondrial translocation of TP53. Interacts with UBD. Interacts with TAF6 isoform 1 and isoform 4. Interacts with C10orf90/FATS; the interaction inhibits binding of TP53 and MDM2 (By similarity). Interacts with NUPR1; interaction is stress-dependent. Forms a complex with EP300 and NUPR1; this complex binds CDKN1A promoter leading to transcriptional induction of CDKN1A. Interacts with PRMT5 in response to DNA damage; the interaction is STRAP dependent. Interacts with PPP1R13L (via SH3 domain and ANK repeats); the interaction inhibits pro-apoptotic activity of p53/TP53. Interacts with PPP1R13B/ASPP1 and TP53BP2/ASPP2; the interactions promotes pro-apototic activity. When phosphorylated at Ser-15, interacts with DDX3X and gamma-tubulin.(Microbial infection) Interacts with cancer-associated/HPV E6 viral proteins leading to ubiquitination and degradation of TP53 giving a possible model for cell growth regulation. This complex formation requires an additional factor, E6-AP, which stably associates with TP53 in the presence of E6.(Microbial infection) Interacts with human cytomegalovirus/HHV-5 protein UL123.(Microbial infection) Interacts (via N-terminus) with human adenovirus 5 E1B-55K protein; this interaction leads to the inhibition of TP53 function and/or its degradation.
Tissue Specificity
Ubiquitous. Isoforms are expressed in a wide range of normal tissues but in a tissue-dependent manner. Isoform 2 is expressed in most normal tissues but is not detected in brain, lung, prostate, muscle, fetal brain, spinal cord and fetal liver. Isoform 3 is expressed in most normal tissues but is not detected in lung, spleen, testis, fetal brain, spinal cord and fetal liver. Isoform 7 is expressed in most normal tissues but is not detected in prostate, uterus, skeletal muscle and breast. Isoform 8 is detected only in colon, bone marrow, testis, fetal brain and intestine. Isoform 9 is expressed in most normal tissues but is not detected in brain, heart, lung, fetal liver, salivary gland, breast or intestine.
Function
Acts as a tumor suppressor in many tumor types; induces growth arrest or apoptosis depending on the physiological circumstances and cell type. Involved in cell cycle regulation as a trans-activator that acts to negatively regulate cell division by controlling a set of genes required for this process. One of the activated genes is an inhibitor of cyclin-dependent kinases. Apoptosis induction seems to be mediated either by stimulation of BAX and FAS antigen expression, or by repression of Bcl-2 expression. Its pro-apoptotic activity is activated via its interaction with PPP1R13B/ASPP1 or TP53BP2/ASPP2. However, this activity is inhibited when the interaction with PPP1R13B/ASPP1 or TP53BP2/ASPP2 is displaced by PPP1R13L/iASPP. In cooperation with mitochondrial PPIF is involved in activating oxidative stress-induced necrosis; the function is largely independent of transcription. Induces the transcription of long intergenic non-coding RNA p21 (lincRNA-p21) and lincRNA-Mkln1. LincRNA-p21 participates in TP53-dependent transcriptional repression leading to apoptosis and seems to have an effect on cell-cycle regulation. Implicated in Notch signaling cross-over. Prevents CDK7 kinase activity when associated to CAK complex in response to DNA damage, thus stopping cell cycle progression. Isoform 2 enhances the transactivation activity of isoform 1 from some but not all TP53-inducible promoters. Isoform 4 suppresses transactivation activity and impairs growth suppression mediated by isoform 1. Isoform 7 inhibits isoform 1-mediated apoptosis. Regulates the circadian clock by repressing CLOCK-ARNTL/BMAL1-mediated transcriptional activation of PER2.
Preparation and Storage
Store at -20 degree C. Stable for 12 months from date of receipt.